

Insome cases of incomplete cervical spinal cord injury (iSCI) there is paresis or weakness of the upper extremity with significantly fewer effects on lower-extremity motor function.1–3Two notable syndromes are associated with these clinical features; the most common is central cord syndrome (CCS)1–6and the second, less-common disorder is known as cruciate paralysis (CP).3–5,7In both syndromes, which are often difficult to differentiate clinically,3the disproportionate effects on the upper limb and hand have been argued to result from selective injury to a somatotopically organized corticospinal tract (CST), damaging fibers controlling the upper limb while sparing those involved in lower-limb movement. In CCS, it has been repeatedly suggested that trauma induces damage to the central gray matter of the cervical spinal cord, which, in turn, selectively damages medially located arm/hand fibers within the lateral CST (LCST) while sparing leg fibers located more laterally within that tract.2,4–6,8As far as we can ascertain, somatotopy or lamination within the LCST was first proposed by Foerster.9However, as Nathan and Smith10noted long ago, Foerster did not provide any supporting evidence to show arm fibers are located medial to leg fibers. In the case of CP, the alleged pathoanatomical mechanism is localized damage to the rostral and medial part of the pyramidal decussation (PD) in the craniovertebral junction (CVJ), where arm fibers first cross the midline, while sparing leg fibers, presumably because they are located laterally and cross the midline caudally. This concept arose from a report by Bell7of high anterior cervical injury causing bilateral upper-extremity paresis with some lower-extremity weakness. Again, no evidence supporting a somatotopically organized decussation was provided. In retrospect, these unsupported anatomical statements,11buttressed by clinical accounts of disproportionate upper- versus lower-limb paresis,7established the idea of an underlying somatotopy at spinal CST levels.
Although it is well established that there exists a somatotopical separation of corticofugal projections from the arm/hand and leg areas of the primary motor cortex (M1) in the corona radiata andinternal capsule,12–15somatotopy of corticospinal fibers as they pass through the CVJ and cervical LCST has been repeatedly challenged.1,3,16,17Early evidence from applying demyelinating axon techniques in postmortem tissue demonstrated that lesions involving M1 arm and hand regions in humans10,18and nonhuman primates (NHPs)12,13,19resulted in degenerating axons scattered throughout the medullary pyramidal tract (PT) and LCST. Similarly, one early tracer study showed overlap between M1 arm and leg fibers within the monkey CVJ.20Furthermore, pathological and neuroimaging evidence in patients with CCS is unclear as to whether the cervical injury site is as restricted as originally suggested by Schneider and colleagues,6or whether it involves more diffuse gray and white matter injury.17,21–23
Another important point is that numerous human and NHP studies have highlighted the relative importance of the cerebral cortex and CST for upper- versus lower-limb function,24–26including the contribution of monosynaptic cortico-motoneuronal connections27–29to the control of hand and finger muscles during skilled grasp and manipulation. As a result, diffuse injury to the tract would generally be expected to lead to a greater deficit in upper- rather than lower-limb function. Despite all this evidence, somatotopy in the LCST remains a common explanation for the disproportionate upper-limb and hand weakness in CCS,2,8,16and remains a sustained feature of teaching in clinical and anatomical textbooks.4,5,16,30,31
To reevaluate the question of somatotopy in the CST, what is needed is a systematic examination of the descending course of corticospinal fibers originating from the arm/hand, shoulder, and leg regions of M1 through the CVJ and cervical spinal cord (C1–T1) in rhesus monkeys, which is the best available model for the human motor system.19,28We have accomplished this by injecting high-resolution anterograde dextran tracers into these regions, which we first identified electrophysiologically. We then used immunohistochemical methods and microscopy to identify labeled axon fibers. Unlike earlier methods, these tracers label both small and large CST fibers, which is important given the enormous preponderance of the former within the CST.11,32
Using an adaptation of the stereological approach,33we also made, for the first time, a quantitative assessment of the location of labeled fibers and were able to demonstrate that there were no significant differences in the relative location within the PT and LCST of the fibers originating from the M1 arm/hand, shoulder, and leg representations. Another important focus of our study was to investigate for the first time the potential somatotopic arrangement of CST fibers from secondary motor areas, including the arm representation of the dorsolateralpremotor cortex(LPMCd), ventrolateral premotor cortex (LPMCv), andsupplementary motor area(M2/SMA). All these secondary areas contribute to the CST, give rise to terminations ending in the cervical spinal gray matter,34–36and contribute to arm and hand movement.28,34Importantly, they may make key contributions to motor recovery after motor system damage.35,37–39
开云体育世界杯赔率
Animals
Eleven adult rhesus macaque monkeys (Macaca mulatta) were used (Table 1).All experimental and neurosurgical procedures were approved by The University of South Dakota Institutional Animal Care and Use Committee and followed United States Department of Agriculture, National Institutes of Health, and Society for Neuroscience guidelines for the ethical treatment of animals.
Description of the parameters in each experimental case
| Case | Gender | Weight (kg) | Area Injected | Tracer/Injections | Total Volume (µl) | Postinjection Survival (days) | Stereological Analysis/Level |
|---|---|---|---|---|---|---|---|
| SDM54 | M | 9.2 | M1 arm/hand | LYD/3 | 1.2 | 33 | PT, C5, C8 |
| SDM57 | F | 6.0 | M1 arm/hand | LYD/3 | 1.2 | 32 | |
| SDM61 | F | 4.3 | M1 arm/hand | LYD/3 | 1.2 | 33 | PT, C5, C8 |
| SDM62 | F | 3.2 | M1 arm/hand | BDA/3 | 1.2 | 33 | |
| IM121 | M | 4.0 | M1 arm/hand | PHA-L/5 | 1.0 | 25 | |
| SDM90 | F | 9.2 | M1 shoulder | BDA/2 | 0.8 | 33 | PT, C5, C8 |
| SDM92 | M | 2.3 | M1 shoulder | BDA/2 | 0.8 | 33 | |
| SDM82 | F | 7.9 | M1 leg | DA488/3 | 1.2 | 33 | PT, C5, C8 |
| SDM84 | M | 11.7 | M1 leg | DA488/2 | 0.8 | 33 | |
| SDM54 | M | 9.2 | LPMCd arm/shoulder | BDA/3 | 1.2 | 33 | PT, C5, C8 |
| SDM57 | F | 6.0 | LPMCd arm/shoulder | FD/3 | 1.2 | 32 | PT,*C5, C8 |
| SDM61 | F | 4.3 | LPMCd arm/shoulder | FD/3 | 1.2 | 33 | |
| SDM72 | F | 5.6 | LPMCd arm/shoulder | BDA/3 | 1.2 | 33 | |
| SDM72 | F | 5.6 | LPMCv arm/shoulder | FD/3 | 1.2 | 33 | |
| SDM54 | M | 9.2 | M2/SMA arm | FD/3 | 1.2 | 33 | PT, C5, C8 |
| SDM62 | F | 3.2 | M2/SMA arm | FD/3 | 1.2 | 33 | |
| SDM77 | M | 9.6 | M2/SMA arm | FD/3 | 1.2 | 33 | PT, C5, C8 |
| SDM84 | M | 11.7 | M2/SMA arm | FD/3 | 1.2 | 33 |
IM = Iowa Monkey; SDM = South Dakota Monkey.
In case SDM57, the brainstem tissue block was cut slightly oblique to the long axis (instead of perpendicular to the long axis as in all other cases), disqualifying the tissue sections from quantitative analysis.
Surgery, Identification of Cortical Areas, and Tracer Injections
所有的神经外科和实验室程序使用to obtain the neuroanatomical data from the cases listed inTable 1have been described previously.35,36,40Briefly, under deep isoflurane anesthesia, a craniotomy was performed to expose the frontomotor cortex. The animal was transferred to ketamine anesthesia and intracortical microsimulation (ICMS) was applied to localize the areas listed inTable 1(M1 arm/hand area, shoulder area, leg area, etc.;Fig. 1).The animal was returned to isoflurane anesthesia and a 0.8- to 1.2-μl injection of biotinylated dextran amine (BDA), lucifer yellow dextran tracer (LYD), fluorescein dextran (FD), phaseolus vulgaris leucoagglutinin (PHA-L), or dextran tracer (DA488) was made into the target region (Table 1).The craniotomy was then closed. Postoperative pain was managed using buprenorphine, and penicillin was used as a pre- and postoperative antibiotic. After a survival period of 25–33 days, the monkey was injected with an intraperitoneal overdose of pentobarbital (50 mg/kg or more) and perfused transcardially using 0.9% saline, followed by a solution of 4% paraformaldehyde in 0.1 M phosphate buffer (PB), 10% sucrose in 0.1 M PB, and then 30% sucrose in PB. The cortex was frozen sectioned at 50 μm in the coronal plane and the brainstem and spinal cord were frozen sectioned at 50 μm in the transverse plane (90° perpendicular to its long axis). In case IM121, the brainstem and spinal cord were sectioned in the coronal/frontal plane. The tissue was processed for immunohistochemical localization of BDA alone, BDA+LYD, BDA+FD, BDA+DA488, or PHA-L alone, as previously described.14,15,36,40

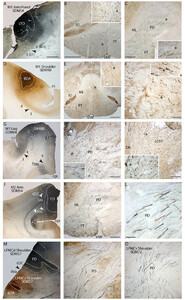
Line drawings of representative cortical injection sites in the primary motor cortex (left column) and premotor cortex (right column).In all cases, high-resolution anterograde tract tracers were injected into a physiologically identified somatotopic representation, mapped using ICMS with current intensities ranging from 1 to 90 μA. For each case, theinsetshows the movement representation map obtained with ICMS. The injection sites are color coded (see legend atbottom left) and thelinewithin each injection site marks the boundary between the centrally located injection site core and the peripherally located injection site halo. Cytoarchitectonic boundaries and cytoarchitectonic regions41are inlight gray。For simplicity, the BDA injection site in case SDM54 (upper left) is not shown. In all cases except SDM72, the plane of section and location for each respective injection site shown inFig. 5(panels A, D, G, J, and M) are identified by thearrowanditalicized panel letter(e.g.,5A).4 = 4区;6 dc =背区域6,尾;6 =博士dorsal area 6, rostral; 6Va = ventral area 6a; 6Vb = ventral area 6b; 6m = area 6 medial; cf = calcarine fissure; cgs = cingulate sulcus; cs = central sulcus; ec = ectocalcarine sulcus; ilas = inferior limb of arcuate sulcus; ios = inferior occipital sulcus; ips = intraparietal sulcus; lf = lateral fissure; ls = lunate sulcus; ots = occipito-temporal sulcus; poms = medial parieto-occipital sulcus; ps = principal sulcus; ros = rostral sulcus; rs = rhinal sulcus; slas = superior limb of the arcuate sulcus; sts = superior temporal sulcus. Movements evoked by ICMS: Ak = ankle; D = digit; El = elbow; L = leg; LL = lower lip; N = neck; NR = no response; Sh = shoulder; T = toe; Th = thumb; Tr = trunk; UL = upper lip; Wr = wrist.
Analysis of Labeling
本地化的皮质注射部位和德xtran-labeled axons through the CVJ and cervical spinal cord (C1–T1) was accomplished using bright-field illumination on a BX-51 Olympus microscope (Leeds Precision Instruments). Attached to the microscope was a high-resolution MAC 5000 motorized stage (Ludl Electronic Products) and Neurolucida neuroanatomical data collection software (MicroBrightField, Inc.) in a Dell Precision Tower 5810 computer. Neurolucida was used to plot major anatomical structures, record cortical injection site locations, and plot the locations of labeled axons through the CVJ and spinal cord using the immunohistochemically stained tissue sections spaced 400–500 μm apart in the CVJ and 1 mm apart in the spinal cord. The cytoarchitectonic location of cortical injections was determined using the Nissl-based criteria and maps of Morecraft et al.36,41Line drawings were created using Adobe Illustrator (Adobe Systems, Inc.;Figs. 1–4).使用显微图像捕获Spotflex 64-Mp shifting pixel camera (Diagnostic Instruments, Inc.), and images were processed using Adobe Photoshop (Adobe Systems, Inc.;Figs. 5and6).
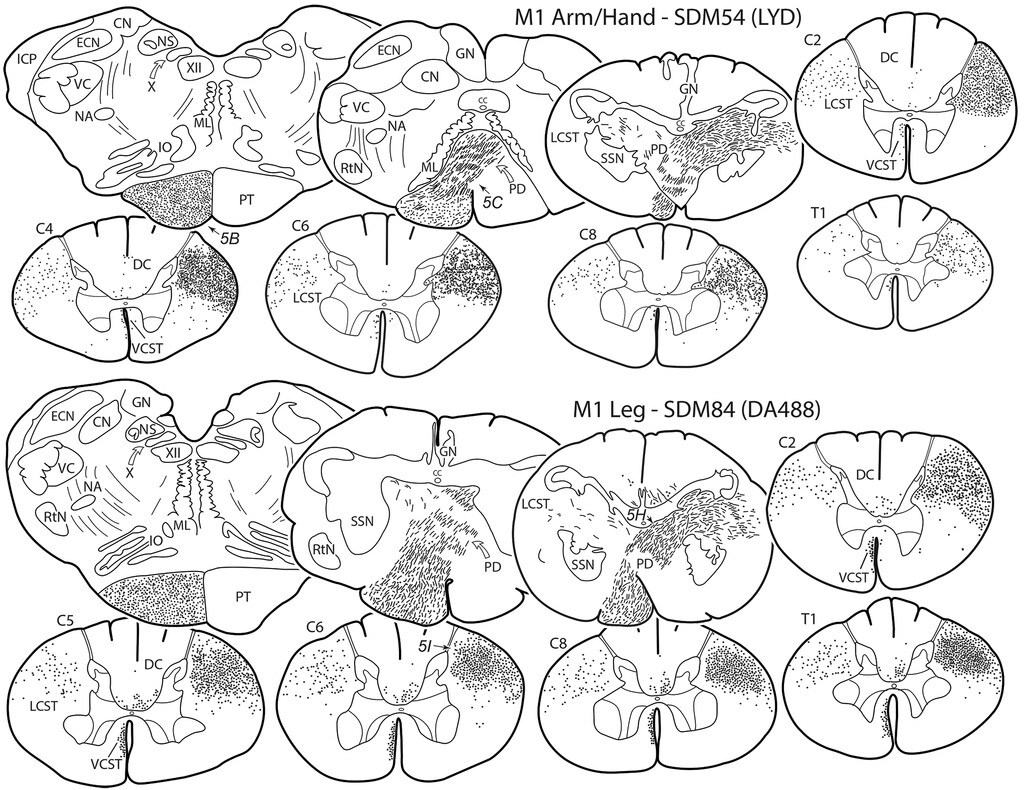
Line drawings depicting selected transverse sections (tissue was cut perpendicular to the long axis of the spinal cord and brainstem) showing the distribution of labeled axons in the CVJ and cervical spinal cord in case SDM54 (upper), which received an injection of LYD in the M1 arm/hand area, and case SDM84 (lower), which received an injection of DA488 in the M1 leg area. For each case, the transverse sections are shown from rostral (upper left, lower medullary pyramid) to caudal (lower right, T1). Note that the locations of photomicrograph panels B, C, and H shown inFig. 5are identified by thearrowanditalicized panel letter(e.g.,5B).CC = central canal; CN = cuneate nucleus; DC = dorsal column; ECN = external cuneate nucleus; GN = gracile nucleus; ICP = inferior cerebellar peduncle; IO = inferior olive; ML = medial lemniscus; NA = nucleus ambiguus; NS = nucleus solitarius; RtN = reticular nucleus; SSN = supraspinal nucleus; VC = trigeminal complex; X = dorsal motor nucleus of the vagus; XII = hypoglossal nucleus.
Line drawings depicting selected transverse sections showing labeled axons in the CVJ and cervical spinal cord in case SDM90 (upper), which received an injection of BDA in the M1 shoulder area, and case SDM54 (lower), which received an injection of FD in the M2/SMA arm area. The transition region between the caudal-most part of the medullary PT and the VCST is indicated by theasteriskin SDM54 (top row, section before C2). For layout of sections and abbreviations, seeFig. 2。请注意,显微照片面板的位置s F and L shown inFig. 5are identified by thearrowanditalicized panel letter(e.g.,5F).
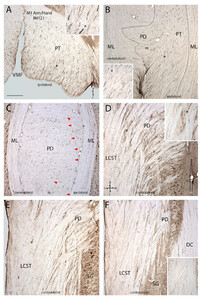
Line drawings depicting selected transverse sections showing labeled axons in the CVJ and cervical spinal cord in case SDM57 (upper), which received an injection of FD in the arm/shoulder area of the LPMCd, and case SDM72 (lower), which received an injection of FD in the arm/shoulder area of the LPMCv. For comparative purposes, the transverse sections for case SDM72 have been reversed (i.e., contralateral is shown on theright).For the layout of sections and standard abbreviations, seeFig. 2。请注意,显微照片面板的位置s N and O shown inFig. 5are identified by thearrowanditalicized panel letter(e.g.,5N).
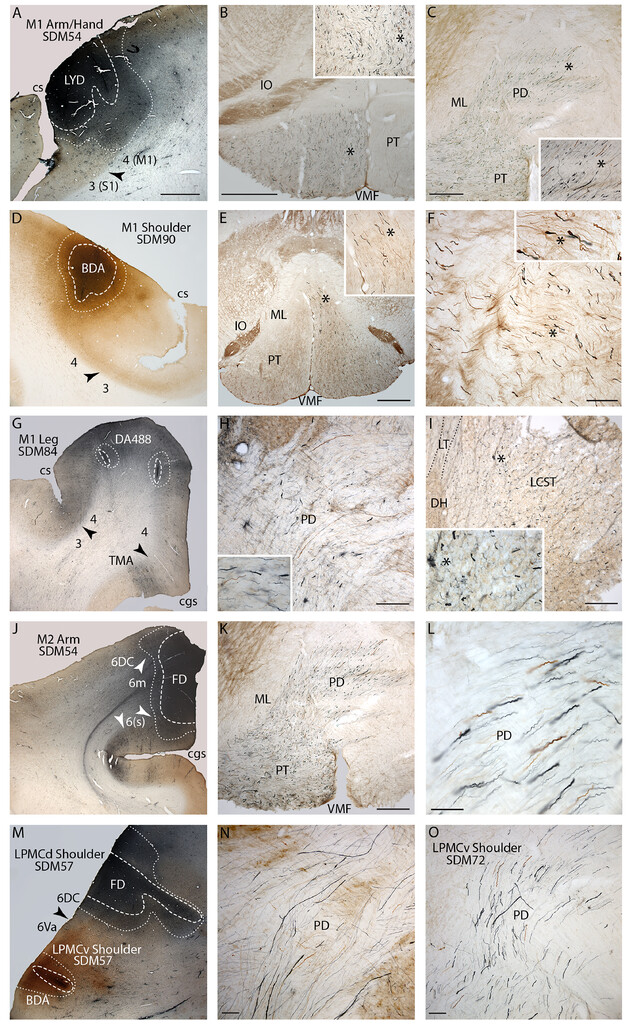
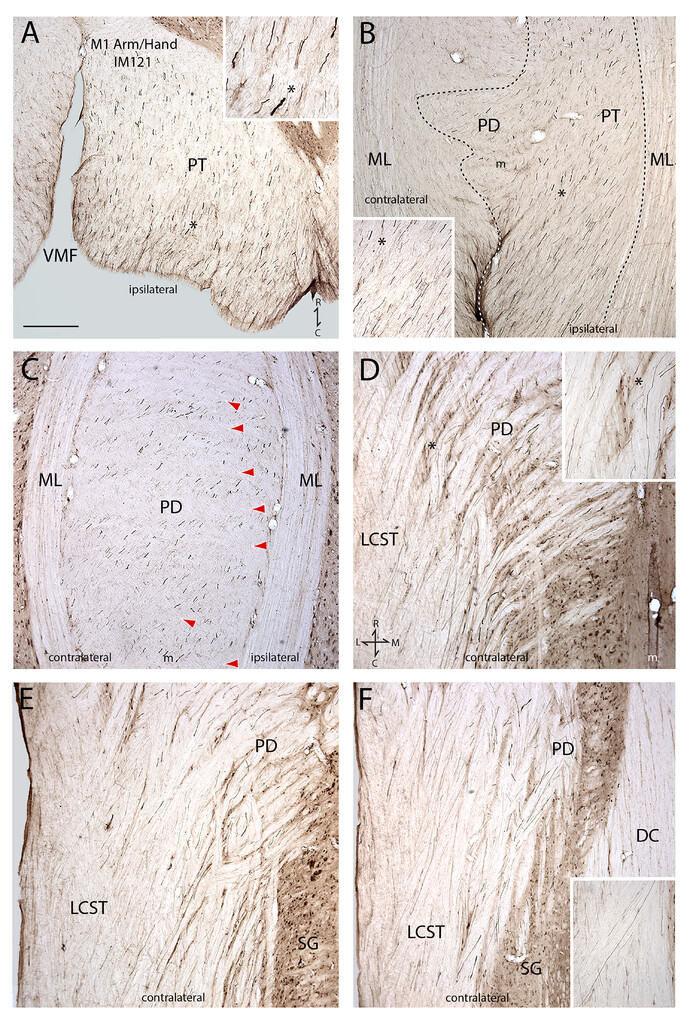
Plate of photomicrographs illustrating representative cortical motor injection sites (coronal plane: panels A, D, G, J, and M) and labeled fibers in the lower medulla and spinal cord (transverse plane: all other panels). All panels from each row are from the same monkey case, except the last row (panels M–O). In all injection sites, the external boundary of the injection site core is identified by thewhite dashed lineand the external boundary of the injection site halo is identified by thewhite dotted line。The core and innermost region of the halo are recognized as the “effective uptake site” where active axonal tracer transport occurs.35,40,43Cytoarchitectonic areas are labeled by Brodmann’s numbers with some contemporary modifications,41and cytoarchitectonic boundaries (seeFig. 1) are identified byblackorwhite arrowheads。注意在每种情况下的核心“吸收地区“was localized to the gray matter, included cytoarchitectonic layer V that contains the somas of CSP neurons, and did not spread into adjacent, unintended cortical motor representations/cytoarchitectonic areas.A:LYD injection site in the central region of the M1 arm/hand area in monkey case SDM54.B: Brainstem section above the PD showing widespread LYD axon labeling (blue) from the M1 arm/hand area in the ipsilateral medullary PT. The intermingledbrown-labeled fibers are from the LPMCd BDA injection in case SDM54. In this and subsequent figures, theasteriskin the main panel marks the location of the higher-power image shown in theinset. C:Section through the upper PD of the CVJ showing LYD-labeled fibers (blue; M1 arm/hand area) andbrown-labeled fibers (LPMCd) leaving the PT and coursing dorsolaterally across the midline.D:BDA injection site in the M1 shoulder area in case SDM90.E:Section through the lower medulla showing widespread BDA-labeled fibers (brown) from the M1 shoulder area passing throughout the ipsilateral medullary PT.F:Section through mid-PD levels showing widespread BDA fiber labeling from the shoulder area of M1.G:DA488 injection site in the M1 leg area in case SDM84.H:DA488-labeled fibers (blue) from the M1 leg area coursing dorsolaterally in the PD. Thebrown-labeled axons are from a BDA injection site placed into another cortical area of interest (M1 face) in case SDM84. Note the complete overlap of M1 leg and M1 face fibers throughout the PD.I:Section through spinal level C6 showing widespread DA488 fiber labeling (blue) from the M1 leg area throughout the cLCST. The dorsal horn gray matter (DH) is identified on the left and Lissauer’s tract (LT) is outlined by dotted lines. As demonstrated by theinset, numerous labeled axons occupy the medial region of the LCST, which according to Foerster9should be label free, but in fact is heavily populated with M1 leg area axons.J:FD injection site in the central region of the M2/SMA arm area in case SDM54.K:Section through lower PT levels, including upper PD levels, showing the widespread distribution of FD-labeled fibers (blue).Thebrown-labeled axons are from the BDA injection site placed in the LPMCd of the same animal and show complete axon overlap from the two secondary motor areas.L:High-power micrograph through mid-PD levels showing complete overlap ofblue(M2 arm area) andbrown(LPMCd) fibers. Note the dominant number of small-diameter fibers scattered among fewer larger-diameter fibers.M:FD injection site in the LPMCd and BDA injection site in the LPMCv in case SDM57.N:Section through mid-/low levels of the PD in case SDM57 showing overlapping FD-labeled axons (blue, LPMCd) and BDA-labeled axons (brown, LPMCd).O:截面上PD水平长效磺胺72 showing FD-labeled fibers (blue) from the LPMCv injection site and BDA-labeled fibers (brown) from the LPMCd injection site. Note that SDM72 injection site photomicrographs are not shown here, but the FD injection in LPMCv is shown in Morecraft et al.36(seeFig. 3D).6(s) = area 6 sulcus; cgs = cingulate sulcus; cs = central sulcus; ML = medial lemniscus; TMA = transitional motor area; VMF = ventral median fissure. Bar = 2 mm (A, D, G, J, and M), 1 mm (B and E), 500 ìm (C and K), 200 ìm (F, H, and I), and 100 ìm (L, N, and O).
Photomicrographs of coronal/frontal sections through the lower medulla and attached upper spinal cord showing the descending course of PHA-L–labeled fibers (black) in case IM121, which received PHA-L injections into the arm/hand representation of M1. The coronal sections are shown progressing from the ventral (A) to dorsal (F) levels of the tissue block. In all panels, as indicated in thebottom rightof panel A, rostral is attopand caudal is atbottom。In photomicrographs with insets, theasteriskdenotes the inset location on the main panel when applicable.A:Coronal section through the midregion of the ipsilateral medullary PT.B:Coronal section through the dorsal region of the PT and beginning (ventral region) of the PD. Thedotted linemarks the outer boundary of the widespread field of labeled M1 arm/hand fibers. Thewhiteportion of thedotted line(to the right of theinset) identifies the ventral (or anterior) median fissure of the medulla.C:Coronal section through the main body (midregion) of the PD. It is very clear that M1 arm/hand fibers cross the midline at all (rostral to caudal) CVJ levels, and not only at rostral levels as suggested by Bell.7Thered arrowheadswithin the decussation show the alternating pattern of unlabeled fiber fascicles (of fibers crossing from the opposite [uninjected] hemisphere) layered between the labeled M1 arm/hand fiber fascicles.D:Coronal section through the dorsal region of the PD and ventral region of the cLCST. Again, note the M1 arm/hand fibers crossing the midline at all rostrocaudal PD levels, and not only at rostral levels, as speculated in CVJ somatotopy theories. Also note here, as well as in panels E and F, that M1 arm/hand fibers occupy medial and lateral regions of the cLCST, and not just the medial region as proposed in Foerster’s theory.9Anatomical orientation is shown in thebottom left。E:Coronal section through midlevels of the cLCST.F:Coronal section through dorsal levels of the cLCST. Theinsetis from a portion of the section below the main panel and shows fibers coursing into the lateral LCST region. Bar = 500 ìm (all main panels). C = caudal; L = lateral; m = midline; M = medial; ML = medial lemniscus; R = rostral; SG = spinal gray matter; VMF = ventral median fissure of the medulla.
Quantification of the Distribution of Labeled Axons in the PT and LCST
Our basic stereological design for quantifying dextran-labeled biological particles has been reported in detail.35,40In the present study, the quantified particle of interest was tracer labeling present within the cross-sections of axons in the PT and LCST (Fig. 7A).Because most of these axons were cut perpendicular to their long axis, each labeled particle represented an individual axon.
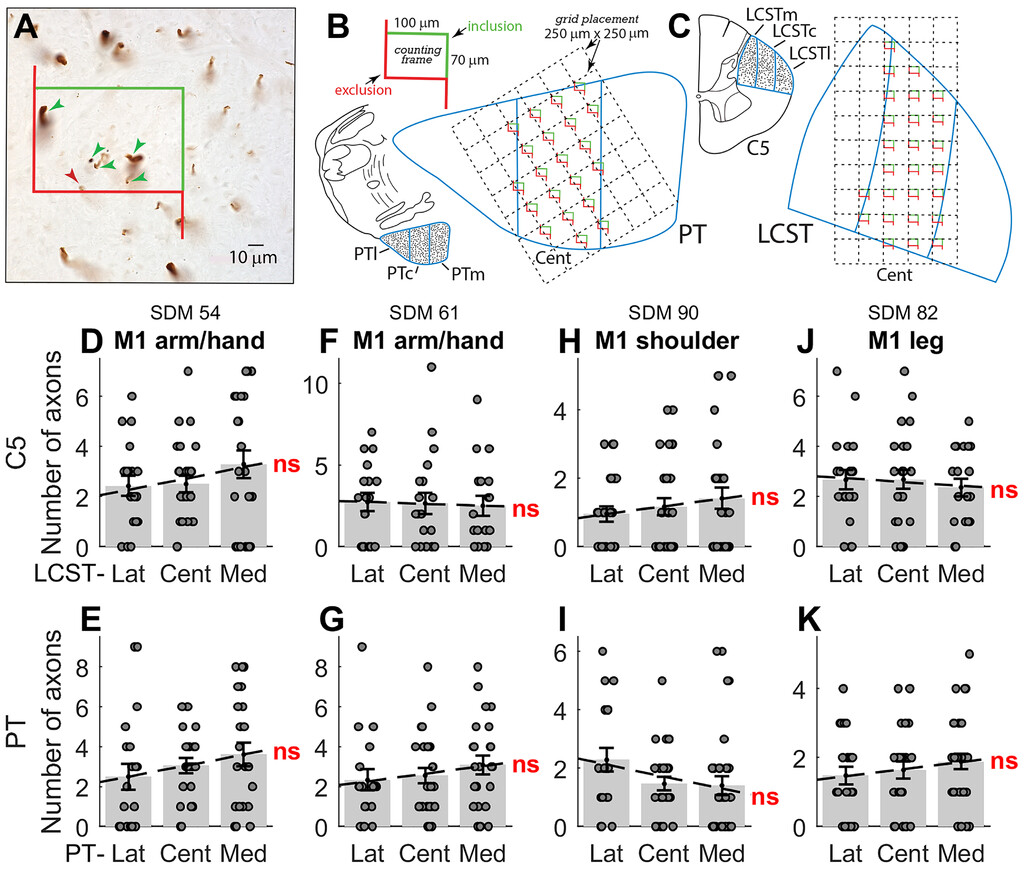
Quantitative assessment of labeled axon numbers within the ROIs of the PT and LCST in representative M1 experimental cases.A:High-power photomicrograph showing an example of a field of BDA-labeled axons in the LCST with an unbiased counting frame randomly placed by the computer software over a sampling area. Labeled axons that were counted in the quantitative analysis included axons located completely within the counting frame (green arrowheads) or axons that were in contact with thegreen “inclusion” line(none in this example). Labeled axons that contacted thered “exclusion” line(red arrowhead) were not counted in the analysis.B:An example of a systematic random placement of the counting grid (250 × 250 μm) and corresponding counting frames placed over the central sector of the medullary pyramidal tract (PTc), just above the PD. A single enlarged counting frame is shown on thetop leftwith the counting frame dimensions (100 × 70 μm) along with thegreen inclusion lineandred exclusion line。C:An example of a systematic random placement of the counting grid (250 × 250 μm) and respective counting frames (100 × 70 μm) placed over the central sector of the LCSTc at C5.D–K:Results from quantitative assessment of labeled fibers in the three subsectors (l, m, and c) in representative C5 (upper row) and PT (lower row) sections for 4 different cases with tracer injection in M1 (D–G = arm/hand; H and I = shoulder; J and K = leg area). Thedotsrepresent the number of fibers within each of the counting frames sampled at random over the entire subsector (B and C). A small random jitter along the x-axis was introduced for visualization purposes only. Thegray barsrepresent the mean (± SEM) number of fibers/counting frame, and thedashed linerepresents the regression calculated across the three subsectors. None of the regressions shown were significant (p = 0.17 [D], 0.16 [E], 0.80 [F], 0.26 [G], 0.24 [H], 0.08 [I], 0.60 [J], and 0.22 [K]). Cent = central; Lat = lateral; LCSTm, LCSTc, and LCSTl = medial, central, and lateral subsectors of the LCST; Med = medial; ns = nonsignificant; PTl, PTc, and PTm = lateral, central, and medial subsectors of the PT.
We adapted Larsen’s stereological approach33to quantify the distribution of labeled axons in the different subsectors of the PT and LCST, using the Optical Fractionator probe in Stereo Investigator 7 (MicroBrightField, Inc.) and a PlanApo ×100 oil objective with a numerical aperture of 1.04 and resolution of 0.2 μm.35,40无偏估计分布的标记axons were obtained using one representative section through the PT immediately above the decussation, one section through mid-C5, and one at mid-C8. The PT and LCST were divided into three equally sized regions of interest (ROIs): lateral, central, and medial.
For counting labeled axons, the main stereological parameters included the counting frame dimensions, counting frame placement grid, and top and bottom guard zones (Fig. 7B and C).The same X/Y counting frame (100 × 70 μm) and X/Y grid placement (250 × 250 μm) were applied to all material. With a random start determined by the software, the unbiased counting frames of area, or a(frame), were systematically placed by precise steps of length (dx and dy) along the x- and y-axes across the entire ROI (e.g., the central subsector of the LCST). The fraction of each ROI area sampled in this study was 12%–15%.
For each ROI we noted the total number of counting frames sampled (14–36, depending on the total ROI area) and the number of labeled axons counted within each frame, following standard exclusion and inclusion counting rules (see Larsen33).We calculated the mean axon count per frame for the different ROIs, and to determine if there was a preferential distribution of labeled axons we performed a 1-way ANOVA with ROI location as a main factor. We also used linear regression to test for any significant mediolateral gradient in the number of labeled fibers across the subsectors of the LCST and confirmed these findings using a robust linear regression analysis (seeSupplemental Table 1).
Results
The injection site in all the cases listed inTable 1was confined to the targeted cortical area and motor representation (Figs. 1, 5A, 5D, 5G, 5J, and 5M).下面,我们就来展示我们的观察的分配ibution of tracer-labeled fibers as they course through the medullary pyramid, PD, LCST, ventral CST (VCST), and dorsal columns (DCs). We also present a brief characterization of terminal labeling in the cervical cord for each corticospinal projection (CSP).
Primary Motor Cortex (M1)
Arm/Hand Area Fibers
Five cases were studied that received injections of a dextran tracer in the M1 arm/hand area (Figs. 1,2,5A–C, and6;Table 1).在所有情况下,注射部位是在to the M1 arm and hand area. In all cases, a dense distribution of fibers occurred throughout the entire ipsilateral pyramid of the medulla. As the labeled fibers approached the rostral part of the PD, most fibers began to arch dorsally and toward the midline in a widely dispersed manner. Once across the midline, the fibers coursed dorsolaterally within the contralateral decussation with no localization. Significantly fewer fibers remained uncrossed and passed into the decussation on the ipsilateral side in a diffuse manner. The heavily labeled contralateral fibers continued throughout the rostral, middle, and caudal parts of the decussation, with no obvious somatotopy. This was evident on all transverse sections through the decussation (Fig. 2),包括IM121脑干和案例spinal cord were sectioned in the coronal/frontal plane (Fig. 6).Thus, axons from the M1 arm/hand area do not selectively lie medially at rostral levels of the decussation as suggested by Bell.3,7In the spinal cord, labeled fibers were densely and widely distributed in the contralateral LCST (cLCST) throughout its medial and lateral parts, with few scattered fibers in the ipsilateral LCST (iLCST). The contralateral projection was dominated by widely distributed and densely packed small- to medium-sized axons with fewer intermingled larger fibers. This pattern was reminiscent of the histopathological accounts of the dispersed LCST distribution of some degenerating large axon fibers among the presence of preserved smaller-diameter fibers in CCS.23,42
At the cervicomedullary junction, a very small contingent of fibers remained in the ventromedial PD region to form the VCST. Most fibers were located ipsilaterally and there was no topography present here either, as labeled fibers were scattered throughout the dorsal, central, and ventral regions of the VCST. In the DC region, 4 of 5 cases had light axon labeling dispersed bilaterally in the ventral DC, near the gray commissural region.
Terminal labeling in segments C1–4 was heavy, primarily targeting the intermediate zone (lamina VII) with some fibers innervating motoneuron lamina IX. A quantitative description of labeling through C5–T1 from this same case material has been published.40Briefly, terminal labeling was also heavy throughout C5–T1, with most (98%) terminations contralateral. Labeling occurred primarily in the contralateral intermediate zone (lamina VII), but a strong contralateral projection occupied motoneuronal lamina IX. Notably, lamina IX terminations gradually increased from C5 to C8/T1, where motoneurons supplying hand and digit muscles are located.
Shoulder Area Fibers
Two cases were examined with an injection in the M1 shoulder area (Figs. 1,3, and5D–F;Table 1).Fiber labeling was heavy and widely distributed throughout the medullary pyramid and contralateral PD, in precisely the same manner as the M1 arm/hand area projection. Labeling in the LCST was also bilateral, widespread, and with a contralateral predominance. In the VCST, both cases gave rise to light bilateral labeling with most fibers located ipsilaterally with no somatotopy. As with the M1 arm/hand projection, labeled fibers were located bilaterally in the ventral DC region.
Significant terminal labeling occurred through the entire cervical spinal cord (C1–T1). The terminal projection was bilateral with a contralateral predominance, primarily involving the intermediate zone (lamina VII). Labeled boutons also occupied the contralateral anterior horn, including motoneuron laminae VIII and IX.
Leg Area Fibers
Two cases were studied with an injection in the M1 leg area (Figs. 1,2, and5G–I;Table 1).In both cases, axon labeling was heavy and widely distributed throughout the lower medullary pyramid and contralateral PD, identical to the pattern found for the M1 arm/hand and shoulder areas. Prominent axon labeling was scattered throughout the rostral, middle, and caudal regions of the decussation at the CVJ. Thus, it was clear that M1 leg axons do not occupy an exclusively lateral position at rostral levels of the decussation, and they do not selectively cross the midline at caudal levels of the decussation as proposed by Bell.7In the LCST, fibers were present bilaterally with a heavy and evenly distributed presence throughout the cLCST (Fig. 5I) with no somatotopy, and a light and scattered distribution in the iLCST. VCST labeling differed in the 2 cases. Only ipsilateral labeling and a moderate number of axons were found in case SDM84 (Fig. 2), whereas light bilateral axon labeling with an ipsilateral predominance occurred in case SDM82. In both cases, there was no clear VCST somatotopy. In both cases, labeled fibers were located in the ventral DC region. In neither case was any terminal labeling found in the cervical spinal gray matter.
Frontal Premotor Cortex (LPMCv, LPMCd, and M2/SMA) Arm/Shoulder Areas
In all premotor cases (Figs. 1,3,4, and5J–O;Table 1), the distribution of axon labeling within the medullary pyramid, PD, and LCST was identical to that for M1 injections. Specifically, labeling was widely distributed in the pyramid, contralateral PD, and cLCST. In the PD, fibers crossed the midline at all rostrocaudal levels. In the LCST, the projection was primarily contralateral with fewer fibers in the iLCST. VCST labeling was absent in many premotor cases. For example, in the LPMCd experiments, VCST axon labeling was bilateral, weak, and predominately ipsilateral in 2 cases and absent in the other 2 cases. In the 1 LPMCv case studied, only weak ipsilateral VCST labeling was found. Finally, VCST labeling in the M2/SMA cases was weak, bilateral in 1 case, ipsilateral in 1 case, and absent in the other 2 cases. DC labeling was absent in some premotor cases. In LPMCd experiments, DC labeling was bilateral, weak, and predominately ipsilateral in 2 cases and absent in the other 2 cases. In the LPMCv case, no DC labeling was found, and in the M2/SMA cases it was bilateral, weak in 2 cases, and absent in the other 2 cases.
In the upper cervical spinal cord (C1–4), the terminal projection from all premotor arm areas was bilateral, primarily involving the contralateral intermediate spinal gray area. Terminal labeling in C5–T1 from the M2/SMA,35LPMCd,36and LPMCv36已经被报道。简而言之,所有情况下了a bilateral projection primarily involving contralateral lamina VII. This CSP was particularly heavy from the M2/SMA arm area (all cases) and caudal LPMCd region (case SDM57 FD). Contralateral terminals were found in motoneuron lamina IX with the M2/SMA primarily innervating the dorsal region and LPMCd the dorsomedial region of lamina IX. The LPMCv did not innervate lamina IX.
Quantitative Analysis of Labeled Fiber Distribution Within the PT and LCST
This analysis was performed on labeled fibers within the PT and within the LCST at C5 and C8 in 8 different cases (Table 1).These cases comprised injections in M1 (arm/hand: 2 cases; shoulder: 1 case; leg: 1 case), in the M2/SMA (arm: 2 cases), and in the LPMCd (arm/shoulder: 2 cases). In each of these cases, single sections from the PT, C5, and C8 were analyzed (except for SDM57 PT;Table 1).Figure 7B and Cshows how the PT and LCST were divided into three subsectors or ROIs (lateral, central, and medial: PTl, PTc, and PTm), and also indicates how the counting frame was moved in a series of steps to allow unbiased samples to be taken across the whole subsector.Figure 7Ashows a high-power photomicrograph of 5 labeled axons within a counting frame, 110 × 70 μm in area.Figure 7D–Kshows the results from 4 M1 injections. In each panel, the number of labeled axons at each site sampled is shown by the gray circles, and the gray bars show the mean number (and SEM) of labeled fibers per sample for each ROI.
The main finding is that in every case (M1 arm [Fig. 7D–G], M1 shoulder [Fig. 7H and I], and M1 leg [Fig. 7J and K]), labeled fibers were present in rather similar numbers in all three subsectors. This was true at all three levels analyzed: PT, C5 (Fig. 7D–K), and C8 (data not shown). The distribution of labeled fibers was broadly similar for the M1 arm/hand (Fig. 7D–G) and leg (Fig. 7J and K) projections. A 1-way ANOVA was performed in each case to determine if the ROI was a significant factor in explaining the variance in fiber distribution across the three ROIs, but in none of the M1 cases did this test return a significant result (Supplemental Table 1).In some cases, there was a trend for more labeled fibers medially than laterally (e.g.,Fig. 7D, E, and K) or vice versa (e.g.,Fig. 7I and J), with very small correlation coefficients (r2, the largest of which was 0.04 inFig. 7I); none of the gradients (indicated by dashed lines inFig. 7D–K) were statistically significant. This was confirmed using a robust linear regression analysis (seeSupplemental Table 1).
Again, the projections from the M2/SMA and LPMCd arm areas also showed similar distributions across three ROIs. In only 1 case (SDM77, arm area of the M2/SMA) was there a significant ANOVA result, and analysis of the gradient indicated a significantly higher number of fibers in the medial subsector than centrally and laterally. However, although this result was confirmed for the PT section by the robust linear regression analysis, this was not the case for the C5 and C8 sections, where the generally very low fiber counts from this injection precluded any secure conclusion from the analysis (Supplemental Table 1).
This quantitative analysis confirmed that there was no clear evidence for somatotopy or lamination in the fiber distribution from different representations within M1 or from all but one of the secondary motor area cases.
Discussion
The results of this investigation firmly reject the concept of somatotopy in the organization of macaque corticospinal fibers as they pass through the PT, CVJ, and cervical LCST. This result is based on the careful identification of fibers at all three levels that were labeled via anterograde transport of modern dextran/lectin tracers injected into different subdivisions (arm/hand, shoulder, and leg) of M1, but also into premotor areas (M2/SMA, LPMCd, and LPMCv). The results were further confirmed by detailed quantitative analysis of the distribution of labeled fibers in the PT and LCST.
In the macaque monkey, an estimated 500,000 fibers in the medullary PT11,32are derived from a number of different cortical areas including, in thefrontal lobe, the M1, supplementary motor area (M2/SMA), and premotor cortex (LPMCd and LPMCv). In most of these areas, but especially in M1, there are clearly defined subregions devoted to the leg, arm/hand, and shoulder. Most of the fibers in the PT continue into the spinal cord, with the majority (approximately 90%) decussating and forming the LCST. The remaining fibers are uncrossed, descending for the most part in the ipsilateral lateral funiculus, while some fibers travel in the VCST.
The present investigation shows that there is overlap in the PT, CVJ, and LCST of fibers anatomically defined as arising from these different cortical areas, and this was confirmed by the quantitative analysis (Fig. 7D–K,Supplemental Table 1).Importantly, from a clinical perspective, which we discuss below, we have also demonstrated that each subpopulation of corticospinal fibers from arm/hand, shoulder, and leg representations within M1 are distributed throughout the territory of the LCST in the cervical cord, and that there is a complete overlap between the distributions of these fiber populations.
It is important to stress that this pattern of intermingling fibers from different cortical motor areas is very different from the spatial separation of corticofugal, including corticospinal, fibers from these areas within the corona radiata, internal capsule, and to some extent within thecerebral peduncle。12–15
Our findings represent a considerable advance over previous results derived from NHP degeneration and horseradish peroxidase tract-tracing studies, which also failed to provide support for somatotopy in the CVJ and cLCST.12,13,20However, our results are far more conclusive because we used powerful high-resolution tract-tracing methodology to account for the contribution of small-diameter fibers (which compose approximately 90% of all PT fibers11,32), detect isolated fiber clusters, and identify individual axons separated from the centralized core of the labeled bundle. These are all important corticospinal axon features that escape detection with the older methods,43are unequivocally resolved in the current study, and enabled our quantitative evaluation (Fig. 7).Indeed, we accurately demonstrate extensive and massive overlap of individual M1 fibers of all diameters at all rostral and caudal levels of the CVJ throughout the entire cross-sectional dimension of the cervicomedullary PT and cLCST (Figs. 2and3).Because this widely distributed pattern also occurred in all premotor cortex experiments (Figs. 3and4), we suggest this represents a fundamental organizational principle of the CST organization as its fibers pass through the decussating PT and LCST. The functional importance of secondary motor areas in recovery from injury is increasingly being recognized.35,37–39
Our findings suggest that potential susceptibility of M1 arm fibers in the VCST20would not explain the disproportionate motor effects following cervical spinal cord injury (SCI) because M1 and premotor arm fiber labeling in the VCST was extremely weak in most cases and absent in others. Pathoanatomical studies show very mild VCST fiber degeneration in patients with CCS.23,42
我们的研究结果表明,轴突内的LCST做not use spatial clues to guide them to their spinal targets. An alternative explanation is that different corticospinal axons destined for control of hand, arm, and leg motoneurons have distinct molecular identities that are expressed early in development as axons enter the decussation/spinal cord. These same mechanisms might explain why there appear to be no terminations put down in the cervical cord by lumbar-heading fibers (seeResults).
Clinical observations in CCS may also reflect a distributed pattern of CST fibers. CCS often has a good prognosis, particularly in younger patients, following selective surgical intervention and rehabilitation.1,4,44Motor recovery is also greater following iSCI compared to complete SCI, and spinal conductivity, in the form of evoked motor potentials, is maintained in spared CSP components of iSCI patients.21,44,45Finally, if leg fibers were to cross through a single, separate (thus vulnerable) region of the caudal CVJ,7one might expect a number of case reports of patients presenting with lower-extremity paralysis/weakness in the absence of upper-limb deficits following lower CVJ/upper cervical injury. But such cases are generally lacking in the literature.
Why Does CCS Have a Greater Impact on Upper- Versus Lower-Limb Function?
This study presents a comprehensive rejection of the concept that there is lamination of corticospinal fibers in the decussation or LCST. It follows that the distinctive impairment of upper-limb (especially hand) function in CCS and in CP cannot be ascribed to focal damage of discretely localized CST fibers destined for upper versus lower limbs, as proposed in many neurological and neurosurgical publications and textbooks.2,4,5,8One possibility is that CCS is largely the result of damage to cervical gray matter,21,45including the motoneurons of arm and hand muscles.46However, if long white matter tracts in the lateral funiculus are also damaged,3,16,21the explanation must now take into account the fact that CST fibers influencing upper-limb movement are widely distributed throughout the decussation and LCST, and are completely intermingled with fibers projecting to the lower limb. Contrary to the opinion originally promulgated by Foerster,9evidence from human neuroanatomy actually argues against lamination of the LCST. Thus, the degeneration within the human LCST following localized infarcts within human M1 is found throughout the tract and is not localized to the medial territory.18
Diffuse injury to the LCST might be expected to produce greater impairment of upper-limb than lower-limb function if CST projections contributed to a much greater extent to the control of upper-limb movements. Several lines of evidence suggest that this is indeed the case. First, it is known that the motor cortical representation of the hand and arm, as defined by electrical stimulation of the cortical surface, is considerably larger than that of the leg and foot. A recent reanalysis of the classic work of Penfield and Boldrey26suggested that comparison of the number of points on the human precentral cortex that produced movement of the arm, hand, or digits was approximately seven times greater than the number of points from which leg, foot, or toe movement was evoked.24Other measures, including precentral cortical surface area or length along thecentral sulcusoccupied by these representations, confirmed the preponderance of upper over lower limb.
Second, while studies using noninvasive stimulation of the human motor cortex report short latency motor evoked potentials in both upper- and lower-limb muscles, these responses are generally larger in the upper versus the lower limbs. For example, there are larger responses in the extrinsic muscles acting on the fingers compared with those acting on the toes.47It is possible that pathways other than the CST are more important for control of lower-limb movements such as locomotion; these pathways may escape damage in CP and CCS.
Finally, it is worth noting that behavioral studies documenting the impact of PT lesions in NHPs are dominated by reports of the effects on the upper limb, and particular skilled hand and digit movements25,48(see Porter and Lemon29).Levi et al.3also noted that larger CST axons—activity that definitely contributes to the earliest excitation of motoneurons supplying hand and digit muscles27–29—are particularly vulnerable to disease and trauma.11,23,42
Technical Considerations
It is important to state that the tracers used in this study are bidirectional, powerful anterograde and moderately retrograde. The evidence shows that retrograde transport does not result in further anterograde transport from noninjected cortical areas.35Indeed, if this were the case, following M1 injections there would have been a powerful CSP to the dorsal horn, which was not found,40because M1 is retrogradely connected with S141and S1, in turn, sends a powerful/massive CSP to the dorsal horn nuclei.49A similar argument can be made for the premotor M2/SMA injections, such that the retrograde interconnection with the caudal cingulate motor area (M4/caudal cingulate motor area [cCMA])41did not result in M2/SMA CSP labeling35in the medial region of the dorsal horn, the primary CSP target of the M4/cCMA.50It is important to stress that injection sites used for this study were confined to the intended somatotopic representation as determined by microscopic analysis of the injection site core and halo regions (Fig. 5A, D, G, J, and M) and ICMS mapping (Fig. 1).Finally, in our quantitative analysis we used a ×100 oil objective with light-level resolution of 0.2 μm, which allowed us to detect nearly all potentially labeled axons because the vast majority of PT axons are greater than 0.25 μm in diameter.32
Conclusions
These results show that within the PD and LCST of the primate there is complete and dense overlap of the territory occupied by M1 leg, shoulder, and arm/hand fibers. This same territory is also occupied by intermingled fibers derived from secondary motor areas of the frontal lobe, including the premotor cortex and SMA. This is the first systematic and quantitative investigation of the distribution—within the cervical spinal cord—of the axons belonging to all these different subdivisions of the CSP. Given the close anatomical parallels between the motor systems of the macaque monkey and humans,19,28,29it seems likely that the symptoms of CCS and CP could reflect the more important contribution of the CST to upper- compared with lower-limb motor function, and not somatotopic organization of the fibers within the tract.
Acknowledgments
We would like to thank Drs. Volker Dietz, Patrick Freund, Armin Curt, and Martin Schubert from the Spinal Cord Injury Centre, University Hospital Zurich, University of Zurich, Switzerland, for providing valuable and constructive input on the paper. We would like to thank Dr. Gary Van Hoesen from the University of Iowa Carver College of Medicine for providing monkey case IM121. This research was supported by NIH grant nos. NS 33003, NS 046367, and NS 097450.
Disclosures
The authors report no conflict of interest concerning the materials or methods used in this study or the findings specified in this paper.
Author Contributions
Conception and design: Morecraft, Lemon. Acquisition of data: Morecraft, Stilwell-Morecraft, Ge. Analysis and interpretation of data: Morecraft, Stilwell-Morecraft, Ge, Lemon, Kraskov. Drafting the article: Morecraft, Stilwell-Morecraft, Ge, Lemon. Critically revising the article: all authors. Reviewed submitted version of manuscript: Morecraft, Stilwell-Morecraft, Ge, Lemon. Approved the final version of the manuscript on behalf of all authors: Morecraft. Statistical analysis: Kraskov. Study supervision: Morecraft.
Supplemental Information
Online-Only Content
Supplemental material is available with the online version of the article.
Supplemental Table 1.//www.prize-show.com/doi/suppl/10.3171/2021.4.JNS202464。
References
-
1 ↑
BadhiwalaJH,AhujaCS,FehlingsMG。Time is spine: a review of translational advances in spinal cord injury. JNSPG 75th Anniversary Invited Review Article。J Neurosurg Spine。2019;30(1):1–18。
-
2 ↑
ChoTA。Spinal cord functional anatomy。Continuum (Minneap Minn)。2015;21(1 Spinal Cord Disorders):13–35。
-
3 ↑
LeviAD,TatorCH,BungeRP。Clinical syndromes associated with disproportionate weakness of the upper versus the lower extremities after cervical spinal cord injury。开云体育app官方网站下载入口。1996;38(1):179–185。
-
5 ↑
RopperA,SamuelsM,KleinJ,PrasadS。Adams and Victors Principles of Neurology。11th ed.McGraw Hill;2019。
-
6 ↑
SchneiderRC,CherryG,PantekH。The syndrome of acute central cervical spinal cord injury; with special reference to the mechanisms involved in hyperextension injuries of cervical spine。J Neurosurg。1954;11(6):546–577。
-
7 ↑
BellHS。瘫痪的双手从伤病上portion of the pyramidal decussation: “cruciate paralysis”。J Neurosurg。1970;33(4):376–380。
-
8 ↑
KunamVK,VelayudhanV,ChaudhryZA,et al.Incomplete cord syndromes: clinical and imaging review。Radiographics。2018;38(4):1201–1222。
-
9 ↑
FoersterO。Symptomatologie der Erkrankungen des Ruckenmarks und Seiner Wurzeln。Springer-Verlag;1936。
-
10 ↑
NathanPW,SmithMC。Long descending tracts in man。I. Review of present knowledge. Brain。1955;78(2):248–303。
-
12 ↑
BarnardJW,WoolseyCN。A study of localization in the cortico-spinal tracts of monkey and rat。J Comp Neurol。1956;105(1):25–50。
-
13 ↑
CoxeWS,LandauWM。Patterns of Marchi degeneration in the monkey pyramidal tract following small discrete cortical lesions。Neurology。1970;20(1):89–100。
-
14 ↑
MorecraftRJ,HerrickJL,Stilwell-MorecraftKS,et al.Localization of arm representation in the corona radiata and internal capsule in the non-human primate。Brain。2002;125(pt 1):176–198。
-
15 ↑
MorecraftRJ,McNealDW,Stilwell-MorecraftKS,et al.Localization of arm representation in the cerebral peduncle of the non-human primate。J Comp Neurol。2007;504(2):149–167。
-
16 ↑
BenglisD,LeviAD。Neurologic findings of craniovertebral junction disease。开云体育app官方网站下载入口。2010;66(3)(suppl):13–21。
-
17 ↑
CollignonF,MartinD,LénelleJ,StevenaertA。Acute traumatic central cord syndrome: magnetic resonance imaging and clinical observations。J Neurosurg。2002;96(1)(suppl):29–33。
-
18 ↑
SchoenJH。Comparative aspects of the descending fibre systems in the spinal cord。Prog Brain Res。1964;11:203–222。
-
19 ↑
KuypersHGJM。Anatomy of the descending pathways。In:BrooksVB, ed.Handbook of Physiology: The Nervous System: Motor Control。Vol II.美国生理学会:1981:597–666。
-
20 ↑
PappasCT,GibsonAR,SonntagVK。Decussation of hind-limb and fore-limb fibers in the monkey corticospinal tract: relevance to cruciate paralysis。J Neurosurg。1991;75(6):935–940。
-
21 ↑
HuberE,DavidG,ThompsonAJ,et al.Dorsal and ventral horn atrophy is associated with clinical outcome after spinal cord injury。Neurology。2018;90(17):e1510–e1522。
-
22
MartinD,SchoenenJ,LenelleJ,et al.MRI-pathological correlations in acute traumatic central cord syndrome: case report。Neuroradiology。1992;34(4):262–266。
-
23 ↑
QuencerRM,BungeRP,EgnorM,et al.Acute traumatic central cord syndrome: MRI-pathological correlations。Neuroradiology。1992;34(2):85–94。
-
25 ↑
LawrenceDG,KuypersHG。The functional organization of the motor system in the monkey. I. The effects of bilateral pyramidal lesions。Brain。1968;91(1):1–14。
-
26 ↑
PenfieldW,BoldreyE。Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation。Brain。1937;60(4):389–443。
-
27
LemonR。Recent advances in our understanding of the primate corticospinal system. F1000 Res。2019;8:F1000。
-
30 ↑
HainesDE,MihaloffGA。Fundamental Neuroscience for Basic and Clinical Applications。5th ed.Elsevier;2018。
-
32 ↑
FirminL,FieldP,MaierMA,et al.Axon diameters and conduction velocities in the macaque pyramidal tract。J Neurophysiol。2014;112(6):1229–1240。
-
34 ↑
MaierMA,ArmandJ,KirkwoodPA,et al.Differences in the corticospinal projection from primary motor cortex and supplementary motor area to macaque upper limb motoneurons: an anatomical and electrophysiological study。Cereb Cortex。2002;12(3):281–296。
-
35 ↑
McNealDW,亲爱的WG,GeJ,et al.Selective long-term reorganization of the corticospinal projection from the supplementary motor cortex following recovery from lateral motor cortex injury。J Comp Neurol。2010;518(5):586–621。
-
36 ↑
MorecraftRJ,GeJ,Stilwell-MorecraftKS,et al.Terminal organization of the corticospinal projection from the lateral premotor cortex to the cervical enlargement (C5-T1) in rhesus monkey。J Comp Neurol。2019;527(16):2761–2789。
-
37
亲爱的WG,GeJ,Stilwell-MorecraftKS,et al.Hand motor recovery following extensive frontoparietal cortical injury is accompanied by upregulated corticoreticular projections in monkey。J Neurosci。2018;38(28):6323–6339。
-
38
KhannaP,TottenD,NovikL,et al.Low-frequency stimulation enhances ensemble co-firing and dexterity after stroke。Cell。2021;184(4):912–930.e20。
-
39
Eisner-JanowiczI,BarbayS,HooverE,et al.Early and late changes in the distal forelimb representation of the supplementary motor area after injury to frontal motor areas in the squirrel monkey。J Neurophysiol。2008;100(3):1498–1512。
-
40 ↑
MorecraftRJ,GeJ,Stilwell-MorecraftKS,et al.Terminal distribution of the corticospinal projection from the hand/arm region of the primary motor cortex to the cervical enlargement in rhesus monkey。J Comp Neurol。2013;521(18):4205–4235。
-
41 ↑
MorecraftRJ,Stilwell-MorecraftKS,CipolloniPB,et al.Cytoarchitecture and cortical connections of the anterior cingulate and adjacent somatomotor fields in the rhesus monkey。Brain Res Bull。2012;87(4-5):457–497。
-
42 ↑
JimenezO,MarcilloA,LeviAD。A histopathological analysis of the human cervical spinal cord in patients with acute traumatic central cord syndrome。Spinal Cord。2000;38(9):532–537。
-
43 ↑
MorecraftRJ,UgolliniG,LanciegoJL,et al.Classic and contemporary neural tract-tracing techniques。In: Johansen-Berg H, Behrens TEJ, eds. Diffusion MRI: From Quantitative Measurement to In Vivo Neuroanatomy。2nd ed.Academic Press;2014:359–399。
-
44 ↑
WirzM,ZörnerB,RuppR,DietzV。Outcome after incomplete spinal cord injury: central cord versus Brown-Sequard syndrome。Spinal Cord。2010;48(5):407–414。
-
45 ↑
CurtA,Van HedelHJ,克劳斯D,DietzV。Recovery from a spinal cord injury: significance of compensation, neural plasticity, and repair。J Neurotrauma。2008;25(6):677–685。
-
46 ↑
GrumblesRM,ThomasCK。Motoneuron death after human spinal cord injury。J Neurotrauma。2017;34(3):581–590。
-
47 ↑
BrouwerB,AshbyP。Corticospinal projections to upper and lower limb spinal motoneurons in man。Electroencephalogr Clin Neurophysiol。1990;76(6):509–519。
-
48 ↑
Hepp-ReymondMC,WiesendangerM。Unilateral pyramidotomy in monkeys: effect on force and speed of a conditioned precision grip。Brain Res。1972;36(1):117–131。
-
49 ↑
Darian-SmithC,LilakA,AlarcónC。Corticospinal sprouting occurs selectively following dorsal rhizotomy in the macaque monkey。J Comp Neurol。2013;521(10):2359–2372。
-
50 ↑
MorecraftRJ,LouieJL,SchroederCM,AvramovK。Segregated parallel inputs to the brachial spinal cord from the cingulate motor cortex in the monkey。Neuroreport。1997;8(18):3933–3938。






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}